Ostrzeżenie naukowców dla ludzkości: mikroorganizmy i zmiany klimatu
Źródło: Nature Reviews Microbiology volume 17, pages 569–586 (2019)

Całe życie na Ziemi, na jednym wykresie. Ile waży całe życie na Ziemi.
Karta Ziemi jako zastępstwo dla Dziesięciu Przykazań
Najgroźniejsze pojedyncze źródło plastiku w oceanach, o którym nikt nie chce mówić – sprzęt rybacki
Streszczenie
W epoce antropocenu, w której obecnie żyjemy, zmiany klimatu wpływają na większość przejawów życia na Ziemi. Mikroorganizmy wspierają istnienie wszystkich wyższych troficznych form życia. Aby zrozumieć w jaki sposób ludzie i inne żywe istoty na Ziemi (łącznie z tymi, których jeszcze nie odkryliśmy) mogą przetrwać antropogeniczne zmiany klimatu, kluczowe jest uwzględnienie wiedzy o „niewidocznej większości”, którą stanowią mikroby. Musimy nauczyć się nie tylko tego jak mikroorganizmy wpływają na zmiany klimatu (łącznie z produkcją i zużyciem gazów cieplarnianych), ale także tego w jaki sposób zmiany klimatu i inne działania człowieka wpływają na zmiany klimatu. Niniejsze Wspólne Oświadczenie dokumentuje centralną rolę i globalne znaczenie zmian klimatu dla biologii. Informuje również ludzkość, że wpływ zmian klimatu będzie w znacznym stopniu zależał od odpowiedzi mikroorganizmów, które są niezbędne, by zapewnić zrównoważoną przyszłość środowiska.
Mikroorganizmy i zmiany klimatu
Wstęp
Działania człowieka oraz ich wpływ na klimat i środowisko doprowadzają do bezprecedensowego wyginięcia zwierząt i roślin, powodują utratę różnorodności biologicznej1,2,3,4 oraz zagrażają życiu zwierząt i roślin na Ziemi5. Przypadki utraty gatunków, społeczności i środowisk życia zostały relatywnie dobrze zbadane, udokumentowane i opisane w publikacjach6. Z mikroorganizmami jest odwrotnie – na ogół nie mówi się o nich w kontekście zmian klimatu (zwłaszcza wpływu zmian klimatu na mikroorganizmy). Chociaż niewidoczne gołym okiem i z tego powodu nieco nieuchwytne7, mnogość (~1030 całkowita liczba bakterii i archeonów)8 i różnorodność mikroorganizmów leżą u podstaw ich roli w utrzymaniu zdrowego światowego ekosystemu: mówiąc najprościej, świat mikrobów stanowi system wsparcia życia biosfery. Choć wpływ człowieka na mikroorganizmy jest mniej oczywisty i z pewnością słabiej scharakteryzowany, najbardziej niepokoi fakt, że zmiany w różnorodności biologicznej i aktywności mikrobów będą miały wpływ na odporność wszystkich innych organizmów i tym samym na ich zdolność odpowiadania na zmiany klimatu9.
Mikroorganizmy odgrywają kluczową rolę w obiegu węgla w przyrodzie i cyklu składników odżywczych, utrzymaniu w zdrowiu zwierząt (w tym ludzi) i roślin, rolnictwie oraz globalnej sieci troficznej. Mikroorganizmy żyją we wszystkich środowiskach na Ziemi zasiedlonych przez organizmy makroskopowe i stanowią jedyne formy życia w innych środowiskach takich jak głęboka podpowierzchnia i środowiska „ekstremalne”. Mikroorganizmy istniały już w na początku życia na Ziemi co najmniej 3,8 miliarda lat temu i prawdopodobne jest, że będą istnieć dużo dłużej niż jakiekolwiek zdarzenia skutkujące wyginięciem gatunków w przyszłości.
Pomimo że mikroorganizmy są kluczowe dla regulacji zmian klimatu, rzadko znajdują się w centrum zainteresowania badań nad zmianami klimatycznymi i nie uwzględnia się ich przy opracowywaniu polityki. Ich olbrzymia różnorodność i zróżnicowane odpowiedzi na zmiany środowiska sprawiają, że określenie ich roli w ekosystemie stanowi wyzwanie. W niniejszym Wspólnym Oświadczeniu ilustrujemy powiązania między mikroorganizmami, organizmami makroskopowymi i zmianami klimatu oraz ostrzegamy ludzkość, że mikroskopijna większość nie może być już dłużej niewidocznym słoniem w pokoju. Jeżeli nie doceniamy znaczenia procesów mikrobiologicznych, w istotnym stopniu ograniczamy nasze rozumienie biosfery Ziemi i odpowiedź na zmiany klimatu, a tym samym narażamy na szwank starania, by stworzyć nieszkodliwą dla środowiska przyszłość6 (Ramka 1).
Ramka 1 Ostrzeżenie naukowców
Sojusz Naukowców Świata i ruch Przestroga Naukowców zostały stworzone, aby ostrzec ludzkość przed wpływem działań człowieka na światowy klimat i środowisko. W 1992 roku 1700 naukowców podpisało pierwsze ostrzeżenie uświadamiając ludziom, że wpływ człowieka naraża przyszłość żyjącego świata na poważne ryzyko267. W 2017 roku, 25 lat później, wystosowano drugie ostrzeżenie w publikacji podpisanej przez ponad 15.000 naukowców5. Ruch ten w dalszym ciągu się rozrasta, a wspomniane ostrzeżenie popiera przeszło 21.000 naukowców. Istotą ostrzeżenia jest wezwanie skierowane do rządów i instytucji, aby w swojej polityce mniej zajmowały się rozwojem gospodarczym, a bardziej ekonomią w zakresie ochrony środowiska, która powstrzyma jego niszczenie i sprawi, że poprzez swoją działalność ludzie zapewnią sobie bezpieczną dla środowiska przyszłość268. Z drugim ostrzeżeniem wiąże się seria artykułów skupiających się na konkretnych tematach, z których pierwszy opisuje znaczenie ochrony bagien269. Celem filmu p.t. „Drugie Ostrzeżenie [The Second Warning]” jest także udokumentowanie poparcia naukowców dla zastąpienia przez ludzkość „zwykłych interesów” i podjęcia działań służących zapewnieniu przetrwania wszystkich gatunków poprzez cofnięcie nieustającego kryzysu związanego ze zmianami środowiska i klimatu.
Cele Sojuszu Naukowców Świata uzupełniają Cele Zrównoważonego Rozwoju Organizacji Narodów Zjednoczonych, które sformułowano, aby wprowadzić w życie zasady godności, pokoju i dobrobytu dla ludzi na całym świecie, teraz i w przyszłości6. Cele te opracowano wokół potrzeb ekonomicznych, społecznych i środowiskowych i odnoszą się one do zrównoważonego rozwoju poprzez eliminację ubóstwa, rozwój bezpiecznych miast i wykształconych społeczeństw, wykorzystywanie odnawialnych źródeł energii (wytwarzanie i zużycie energii) oraz niecierpiące zwłoki działania dotyczące zmian klimatycznych uwzględniające sprawiedliwe wykorzystywanie systemów wodnych i naziemnych w celu uzyskania zdrowej, mniej zanieczyszczonej biosfery. Cele te uwzględniają, że odpowiedzialne zarządzanie ograniczonymi zasobami naturalnymi jest konieczne dla rozwoju odpornych[rezylientnych], stabilnych społeczeństw.
Nasze Wspólne Oświadczenie stanowi ostrzeżenie dla ludzkości z punktu widzenia mikrobiologii. Jako że jest to ostrzeżenie mikrobiologów, stawia sobie za cel zwiększenie świadomości dotyczącej świata mikrobów oraz wezwanie do działania dla mikrobiologów, aby coraz bardziej angażowali się w i na rzecz badań nad drobnoustrojami, tak aby stawały się one coraz większą częścią programów ramowych dotyczących zmian klimatycznych i aby możliwe było osiągnięcie Celów Zrównoważonego Rozwoju Organizacji Narodów Zjednoczonych (Ramka 2). Bazuje na wcześniejszych wysiłkach naukowców i w zakresie polityki mających na celu zwrócenie uwagi na rolę mikroorganizmów w zmianach klimatu7,126,270,271,272 i na ich istotne znaczenie dla społeczeństwa7. Mikrobiolodzy są w stanie poprzeć ostrzeżenie mikrobiologów stając się sygnatariuszami.
Zakres Wspólnego Oświadczenia
W niniejszym Wspólnym Oświadczeniu poruszamy temat wpływu mikroorganizmów na zmiany klimatu, w tym mikrobiologicznych procesów aktywnych klimatycznie i ich siły napędowej. Odnosimy się również do wpływu zmian klimatycznych na drobnoustroje skupiając się na oddziaływaniu zmian klimatu na skład i funkcjonowanie środowiska drobnoustrojów, reakcjach fizjologicznych oraz adaptacji na drodze ewolucji. Mimo że w centrum naszego zainteresowania znajdują się powiązania między mikroorganizmami a klimatem, rozważamy także działalność człowieka wywierającą mniej bezpośredni, ale niewykluczone, że synergistyczny wpływ, jak np. lokalne zanieczyszczenie czy eutrofizacja.
Dla celów niniejszego Wspólnego Oświadczenia definiujemy „mikroorganizm” jako dowolny mikroskopijny organizm lub wirus niewidoczny gołym okiem (mniejszy niż 50 µm), który może istnieć w postaci jednokomórkowej, wielokomórkowej (na przykład gatunki różnicujące), zagregowanej czy też w postaci wirusa. Oprócz mikroskopijnych bakterii, archeonów, eukariontów i wirusów, omawiamy także pewne makroskopijne eukarionty jednokomórkowe (na przykład rozległy morski fitoplankton) oraz grzyby powodujące butwienie drewna. Naszym zamiarem nie jest wyczerpujące omówienie wszystkich środowisk ani wszystkich wpływów antropogenicznych, lecz przedstawienie przykładów z głównych globalnych biomów (morskich i lądowych), które uwydatniają wpływ zmian klimatu na procesy mikrobiologiczne i ich skutki. Zwracamy również uwagę na rolnictwo i choroby zakaźne oraz na rolę mikroorganizmów w łagodzeniu zmian klimatycznych. Nasze Wspólne Oświadczenie alarmuje mikrobiologów i innych naukowców o konieczności zajęcia się tematem roli mikroorganizmów w nasilaniu lub minimalizowaniu wpływu antropogenicznych zmian klimatu (Ramka 1).
Biom morski
Biomy morskie zajmują ~70% powierzchni Ziemi i obejmują zarówno ujścia rzek do morza, jak też namorzyny, rafy koralowe i otwarte oceany (Ilustracja 1). Mikroorganizmy fototropiczne wykorzystują energię ze słońca w górnych 200 m kolumny wodnej, podczas gdy organizmy morskie w głębszych strefach czerpią energię z organicznych i nieorganicznych substancji chemicznych10. Poza światłem słonecznym, dostępność innych postaci energii oraz temperatura wody (wahająca się od mniej więcej -2°C na morzach pokrytych lodem do ponad 100°C w kominach hydrotermalnych) ma również wpływ na strukturę środowiska morskiego11. Rosnące temperatury nie tylko wpływają na procesy biologiczne, ale także zmniejszają gęstość wody i przez to również stratyfikację i cyrkulację, co ma wpływ na rozprzestrzenienie się organizmów i transport składników pokarmowych. Opady atmosferyczne, zasolenie i wiatry także wpływają na stratyfikację, mieszanie i cyrkulację. Napływ składników pokarmowych z powietrza, rzek i ujść rzek do morza również oddziałuje na strukturę i funkcjonowanie społeczności mikrobiologicznej, zaś zmiany klimatu oddziałują na wszystkie te czynniki fizyczne.
Ilustracja 1: Mikroorganizmy i zmiany klimatyczne w biomach morskich i lądowych.
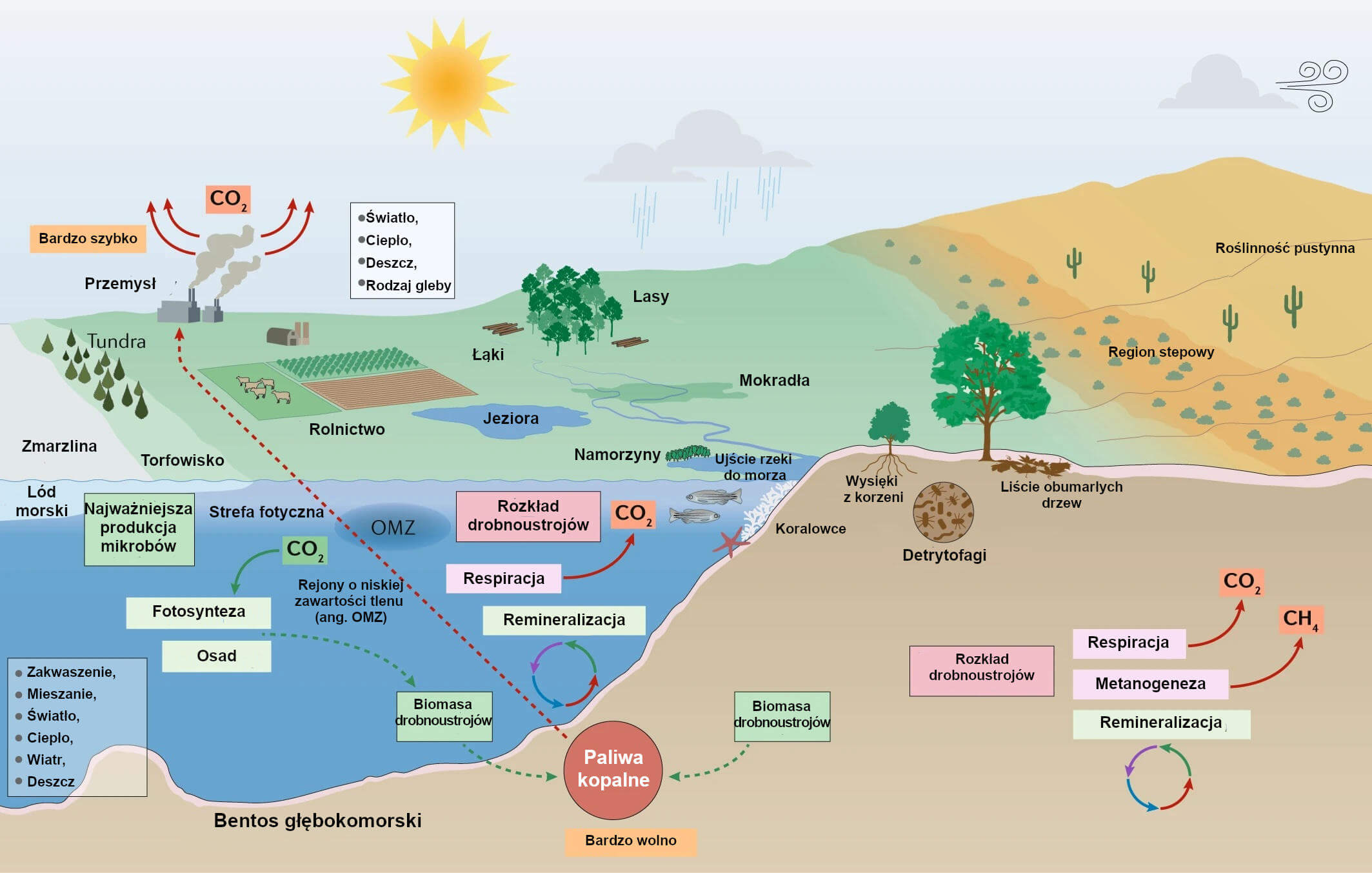
W środowisku morskim najważniejsza produkcja mikrobów znacząco przyczynia się do sekwestracji CO2. Mikroorganizmy morskie utylizują ponadto składniki pokarmowe wykorzystywane w morskiej sieci troficznej i w trakcie tego procesu uwalniają CO2 do atmosfery. W wielu różnych środowiskach lądowych mikroorganizmy są kluczowymi reducentami materii organicznej i uwalniają do atmosfery składniki pokarmowe w glebie, jak też CO2 i CH4, umożliwiając wzrost roślin. Biomasa drobnoustrojów i inna materia organiczna (pozostałości roślin i zwierząt) jest przetwarzana w paliwa kopalne w ciągu milionów lat. Spalanie paliw kopalnych, przeciwnie, uwalnia gazy cieplarniane przez niewielki ułamek tego czasu. W rezultacie obieg węgla w przyrodzie jest skrajnie zaburzony, zaś poziomy CO2 w atmosferze będą nadal rosły tak długo jak będzie się spalać paliwa kopalne. Liczne przejawy działalności człowieka, w tym rolnictwo, przemysł, transport, wzrost liczby ludności i konsumpcji przez człowieka w połączeniu z lokalnymi czynnikami środowiskowymi takimi jak rodzaj gleby i światło, wywierają znaczny wpływ na skomplikowaną sieć interakcji drobnoustrojów z innymi mikroorganizmami, roślinami i zwierzętami. Interakcje te dyktują w jaki sposób mikroorganizmy reagują na zmiany klimatu i na nie wpływają (na przykład poprzez emisjię gazów cieplarnianych), a także jak zmiany klimatyczne (na przykład wyższe poziomy CO2, ocieplenie i zmiany w opadach atmosferycznych) wpływają z kolei na reakcje mikrobiologiczne. OMZ, rejony o niskiej zawartości tlenu.
Ogólne znaczenie mikroorganizmów dla ekosystemów oceanów można docenić biorąc pod uwagę ich liczbę i biomasę w kolumnie wodnej i podpowierzchni: całkowita liczba komórek przekracza 1029 (źródła 8,12,13,14,15,16), zaś Spis morskich form życia szacuje, że 90% morskiej biomasy stanowią drobnoustroje. Poza tymi suchymi liczbami, mikroorganizmy morskie pełnią kluczowe funkcje dla ekosystemu. Wiążąc do postaci stałej węgiel i azot oraz remineralizując materię organiczną, mikroorganizmy morskie tworzą podstawę sieci troficznych oceanów, a tym samym globalny obieg węgla i składników pokarmowych w przyrodzie13. Osiadanie, odkładanie się i spalanie węgla stałego w drobnocząsteczkowej materii organicznej do morskich osadów stanowi kluczowy długofalowy mechanizm sekwestracji CO2 z atmosfery. Z tego powodu równowaga pomiędzy regeneracją CO2 i składnikami pokarmowymi poprzez remineralizację w porównaniu z zagrzebaniem na dnie morza determinuje wpływ na zmiany klimatu.
Poza ociepleniem (na skutek rosnących stężeń CO2 w atmosferze potęgujących efekt cieplarniany) zakwaszenie oceanów wzrosło o ~0.1 jednostek pH od czasów przedindustrialnych, przy czym przewiduje się, że do końca tego wieku nastąpią dalsze redukcje wynoszące 0.3-0.4 jednostek17,18,18. Biorąc pod uwagę bezprecedensowe tempo zmiany pH19,20,21, istnieje potrzeba szybkiego uzyskania wiedzy w jaki sposób zareaguje życie w morzu22. Wpływ zwiększonych stężeń gazów cieplarnianych na temperaturę oceanów, zakwaszenie, stratyfikację, mieszanie, cyrkulację termohalinową, zasoby składników pokarmowych, światło oraz ekstremalne zdarzenia pogodowe oddziałuje na mikroflorę morską na wiele różnych sposobów, które niosą istotne konsekwencje dla środowiska, w tym poważne zmiany produktywności, morskiej sieci troficznej, eksporcie węgla i zagrzebywaniu na dnie morza19,23,24,25,26,27,28,29.
Mikroorganizmy wpływają na zmiany klimatu
Fitoplankton morski odpowiedzialny jest za połowę światowego fotosyntetycznego wiązania CO2 (najważniejsza globalna produkcja netto wynosząca ~50 Pg węgla rocznie) i za połowę wytwarzania tlenu pomimo tego, że wynosi zaledwie ~1% światowej biomasy roślinnej30. W porównaniu z roślinami lądowymi fitoplankton morski rozproszony jest na bardziej rozległym obszarze powierzchni wody, narażony jest na mniejszą sezonową zmienność i ma wyraźnie szybsze tempo odnowy niż drzewa (dni w porównaniu do dekad)30. Z tych powodów fitoplankton w skali globalnej reaguje szybko na zmienność klimatu. Cechy te są ważne, kiedy ocenia się wkład fitoplanktonu w wiązania węgla i przewiduje w jaki sposób produkcja ta może się zmienić w odpowiedzi na zakłócenia. Prognozy dotyczące wpływu zmian klimatycznych na najważniejszą produktywność komplikują cykle kwitnienia fitoplanktonu, na które wpływa zarówno kontrola z dołu do góry (na przykład dostępność niezbędnych składników pokarmowych i mieszanie pionowe), jak i z góry do dołu (na przykład pastwiska i wirusy)27,30,31,32,33,34. Nasilenie promieniowania słonecznego, temperatury i dopływów słodkiej wody do wód powierzchniowych wzmacniają stratyfikację oceanów i w efekcie redukują transport składników pokarmowych z wód głębinowych do powierzchniowych, co zmniejsza najważniejszą produktywność. Wzrost poziomów CO2, odwrotnie, może zwiększyć najważniejszą produkcję fitoplanktonu, ale tylko wtedy, gdy nie brakuje składników pokarmowych36,37,38.
Niektóre badania wskazują, że ogólne światowe zagęszczenie fitoplanktonu oceanicznego zmniejszyło się w ostatnim stuleciu39, ale wnioski te kwestionuje się ze względu na ograniczoną dostępność długofalowych danych dotyczących fitoplanktonu, różnice metodologiczne w generowaniu danych oraz znaczne zróżnicowanie produkcji fitoplanktonu40,41,42,43. Co więcej, inne badania sugerują globalny wzrost produkcji fitoplanktonu44 i zmiany w konkretnych regionach lub w określonych grupach fitoplanktonu45,46. Globalny lód morski (patrz Indeks Lodu Morskiego) zanika, co prowadzi do wyższej przepuszczalności światła i potencjalnie zwiększonej najważniejszej produkcji47; istnieją jednak wykluczające się prognozy na temat wpływu zróżnicowanych wzorców mieszania i zmian w zasobach środków pokarmowych oraz tendencji dotyczących produktywności w strefach polarnych34. Podkreśla to potrzebę zgromadzenia długofalowych danych dotyczących produkcji fitoplanktonu oraz składu społeczności drobnoustrojów. Długofalowe dane potrzebne są, aby rzetelnie przewidzieć w jaki sposób funkcje mikrobów i mechanizmy sprzężenia zwrotnego zareagują na zmiany klimatu, a jednak istnieje bardzo niewiele takich właśnie zbiorów danych (na przykład badania Hawaii Ocean Time-series i Bermuda Atlantic Time-series)48,49,50. W tym kontekście Global Ocean Sampling Expedition51, transekty Oceanu Południowego52,53, a także Tara Oceans Consortium11,54,55,56,57,58,59 dostarczają dane metagenomiczne, które stanowią cenny punkt odniesienia dla mikroorganizmów morskich.
Okrzemki są odpowiedzialne za 25-45% całkowitej najważniejszej produkcji w oceanach60,61,62 dzięki temu, że występują powszechnie w rejonach otwartych oceanów, kiedy całkowita biomasa fitoplanktonu jest maksymalna63. Okrzemki mają relatywnie dużą prędkość tonięcia w porównaniu z innymi grupami fitoplanktonu i odpowiadają za ~40% eksportu węgla drobnocząsteczkowego do głębin62,64. Fizycznie napędzane sezonowe wzbogacenia składników pokarmowych na powierzchni sprzyjają kwitnieniu okrzemków. Antropogeniczne zmiany klimatu bezpośrednio wpłyną na te cykle sezonowe zmieniając czas kwitnienia i zmniejszając ich biomasę, co zredukuje ich najważniejszą produkcję i absorpcję CO265. Dane teledetekcji wskazują na zmniejszenie się liczebności okrzemków w skali światowej między 1998 a 2012 rokiem, zwłaszcza na Północnym Pacyfiku, co wiąże się z płyceniem powierzchniowej warstwy dobrze wymieszanej oraz z niższymi stężeniami składników pokarmowych46.
Oprócz wkładu fitoplanktonu morskiego w sekwestrację CO230,66,67,68, archeony chemolitotroficzne i bakterie wiążą CO2 w ciemnościach panujących w głębokich wodach oceanów69 i na powierzchni w okresie zimy polarnej70. Bakterie morskie i archeony przyczyniają się także w istotnym stopniu do respiracji powierzchni oceanów oraz krążenia wielu elementów składowych18. Metanogeny i metanotrofy na dnie oceanów są ważnymi producentami i konsumentami CH4, ale ich wpływ na strumień atmosferyczny tego gazu cieplarnianego jest niepewny71. Wirusy morskie, bakterie bdellovibrio i eukariotyczne organizmy żerujące są również istotnymi elementami składowymi mikrobiologicznej sieci troficznej; na przykład, wirusy morskie oddziałują na skuteczność sekwestracji węgla i jego odkładania się w głębinach oceanów57. Zmiany klimatu wpływają na interakcje pomiędzy drapieżnikami a ofiarami, w tym na wzajemne oddziaływanie wirusów i żywicieli, a przez to na globalne cykle biogeochemiczne72.
Rejony o niskiej zawartości tlenu (OMZs) zwiększyły swoją powierzchnię w ciągu ostatnich 50 lat na skutek ocieplania się oceanów, które redukuje rozpuszczalność tlenu73,74,75. OMZs pełnią rolę światowych zbiorników wodnych wysychających dla wchodzącego w reakcje azotu, zaś produkcja N2 i N2O przez drobnoustroje odpowiada za ~25-50% utraty azotu z oceanów do atmosfery. Ponadto OMZs stanowią najbardziej rozległe morskie rezerwuary metanu w oceanach i znacząco przyczyniają się do cyklów metanu na otwartym oceanie. Z tych przyczyn obserwowana i przewidywana przyszła ekspansja rejonów o niskiej zawartości tlenu (OMZs) może istotnie wpłynąć na bilans składników pokarmowych i gazów cieplarnianych, a także na rozmieszczenie organizmów zależnych od tlenu73,74,75.
Górne 50 cm osadu głębokiego morza składa się z ~1 x 1029 mikroorganizmów8,16, zaś całkowite zagęszczenie archeonów i bakterii w tych osadach zwiększa się wraz z szerokością geograficzną (od 34°N do 79°N), przy czym konkretne jednostki taksonomiczne przyczyniają się nieproporcjonalnie do tego wzrostu76. Mikroorganizmy zasiedlające strefę bentoniczną wykazują wzorce biogeograficzne i reagują na zmiany ilości oraz jakości materii cząsteczkowej opadającej na dno oceanu77. W efekcie naukowcy spodziewają się, że zmiany klimatu w szczególności dotkną procesy funkcjonalne, które wykonują archeony strefy bentonicznej zasiedlające głębokie morze (takie jak utlenianie amoniaku) i związane z nimi cykle biogeochemiczne76.
Aerozole mają wpływ na powstawanie chmur, przez co oddziałują też na światło słoneczne i na opady, ale stopień w jakim wpływają na zmiany klimatyczne i sposób, w jaki to czynią pozostają nieokreślone78. Aerozole morskie składają się ze skomplikowanej mieszanki soli morskiej, siarczanu nie pochodzącego z soli morskiej oraz cząsteczek organicznych i mogą funkcjonować jako jądra kondensacji wpływając na równowagę promieniowania i z tego powodu również na klimat79,80. Dla przykładu aerozole biogeniczne w odległych środowiskach morskich (na przykład na Oceanie Południowym) mogą zwiększać liczbę i rozmiar kropli chmurowych oddziałując na klimat w podobny sposób, jak aerozole w rejonach o wysokim skażeniu80,81,82,83. W szczególności fitoplankton emituje siarczek dimetylu, zaś jego pochodny siarczan pobudza kondensację chmur79,84. Zrozumienie sposobów, w jakie fitoplankton morski przyczynia się do aerozolów pozwoli lepiej przewidzieć jak zmieniające się warunki w oceanie wpłyną na chmury i przejdą w sprzężenie zwrotne wobec klimatu84. Oprócz tego sama atmosfera zawiera ~1022 komórek mikrobów, zaś określenie zdolności mikroorganizmów żyjących w atmosferze do wzrostu i tworzenia grup będzie miało wartość przy ocenie ich wpływu na klimat8.
Porośnięte roślinnością środowiska są istotne dla sekwestracji węgla określonej przez całą gamę troficzną od drapieżników po zwierzęta roślinożerne oraz rośliny i związane z nimi społeczności drobnoustrojów85. Działalność człowieka, w tym antropogeniczne zmiany klimatu, zredukowała te środowiska w ciągu ostatnich 50 lat o 25-50%, natomiast całkowita liczba morskich drapieżników spadła nawet o 90%85,86,87. Biorąc pod uwagę tak znaczne zakłócenia, należy ocenić wpływ na społeczności mikrobów, ponieważ aktywność drobnoustrojów determinuje ilość węgla, jaka ulega remineralizacji i uwalnia się jako CO2 i CH4.
Zmiany klimatu wpływają na mikroorganizmy
Zmiany klimatyczne zaburzają wzajemne oddziaływanie między gatunkami i zmuszają gatunki, by się przystosowywały, migrowały i były zastępowane przez inne lub wyginęły28,88. Wzrost temperatury oceanów, zakwaszenie, eutrofizacja i nadmierne wykorzystywanie (na przykład rybołówstwo, turystyka) łącznie powodują niszczenie raf koralowych i mogą wywoływać przesuwanie się ekosystemów w stronę wodorostów89,90,91,92,93 oraz bentonicznych mat sinic94,95. Na zdolność koralowców do przystosowywania się do zmian klimatu w znacznym stopniu wpływają reakcje powiązanych z nimi mikroorganizmów, w tym mikroalgalnych symbiontów i bakterii96,97,98. Od setek do tysięcy gatunków mikrobów żyjących na koralowcach jest kluczowych dla zdrowia żywiciela, na przykład poprzez utylizowanie zbytecznych produktów przemiany materii, zapewnianie niezbędnych składników pokarmowych i witamin oraz wspieranie układu odpornościowego w walce z patogenami99. Jednakże zakłócenia w środowisku lub utrata koloru przez koralowce mogą szybko zmienić mikrobiom koralowca. Zmiany takie niewątpliwie wpływają na funkcje ekologiczne i stabilność systemu koralowiec-mikroorganizm potencjalnie oddziałując na zdolność i tempo, w jakim koralowce przystosowują się do zmian klimatycznych, jak również relacje między koralowcami a innymi częściami składowymi ekosystemu rafy99,100.
Ogólnie rzecz biorąc mikroorganizmy mogą rozprzestrzenić się łatwiej niż organizmy makroskopijne. Niemniej jednak u wielu gatunków mikrobiologicznych występują różnice biogeograficzne, zaś na skład i funkcjonowanie społeczności silny wpływ wywierają czynniki związane z rozmieszczeniem, stylem życia (na przykład związek z żywicielem) oraz czynniki środowiskowe54,101,102,103. Prądy oceaniczne oraz gradienty temperatury i gradienty równoleżnikowe są szczególnie istotne dla społeczności morskich104,105. Jeśli przemieszczenie się do bardziej sprzyjających środowisk nie jest możliwe, zmiana ewolucyjna może być jedynym mechanizmem przetrwania88. Mikroorganizmy takie jak bakterie, archeony i mikroalgi, charakteryzujące się dużą liczebnością i szybkim cyklem rozmnażania bezpłciowego posiadają wysoki potencjał adaptacyjny22. W relatywnie niewielu badaniach przeanalizowano adaptację ewolucyjną do zakwaszenia oceanów lub innych zmiennych środowiskowych odnoszących się do zmian klimatu22,28. Analogicznie, w ograniczonym stopniu rozumiemy mechanizmy cząsteczkowe reakcji fizjologicznych i konsekwencji tych reakcji dla cyklów biogeochemicznych18.
Jednakże w kilku badaniach wykazano wpływ podwyższonych poziomów CO2 na poszczególne gatunki fitoplanktonu, który może zakłócać bardziej ogólne procesy na poziomie ekosystemu. Eksperyment w terenie udowodnił, że rosnące poziomy CO2 zapewniają selektywną korzyść toksycznym mikroalgom Vicicitus globosus, prowadząc do zaburzeń transportu materii organicznej przez poziomy troficzne106. Rodzaj morskich sinic Trichodesmium reaguje na długotrwałą (4,5 lat) ekspozycję na podwyższone poziomy CO2 nieodwracalnymi zmianami genetycznymi, które zwiększają wiązania azotu i jego poziom107. W przypadku fotosyntetycznej zielonej algi Ostreococcus tauri, podwyższone poziomy CO2 przyśpieszają wzrost, rozmiar komórek i stosunek węgla do azotu108. Wyższe poziomy CO2 wpływają także na strukturę populacji O. tauri prowadząc do zmian ekotypów i zasiedlania niszowego i tym samym oddziałując na szersze sieci troficzne oraz cykle biogeochemiczne108.
Zamiast wytwarzać większe komórki, wapniejące gatunki fitoplanktonu Emiliania huxleyi reagują na łączny wpływ podwyższonej temperatury i podwyższonych poziomów CO2 (i związanego z nimi zakwaszenia) produkując mniejsze komórki, które zawierają mniej węgla109. Jednakże dla tego gatunku ogólne wskaźniki produkcji nie zmieniają się na skutek ewolucyjnej adaptacji do wyższych poziomów CO2109. Reakcje na poziomy CO2 różnią się dla poszczególnych społeczności (na przykład między fitoplanktonem w Arktyce a fitoplanktonem na Antarktydzie110). Badanie mezokosmosu rozpoznało modyfikacje zmiennych w różnorodności wirusów, które infekują E. huxleyi, kiedy rośnie w otoczeniu podwyższonych poziomów CO2, jak również zwróciło uwagę na potrzebę ustalenia czy podwyższone poziomy CO2 wpływają bezpośrednio na wirusy, żywicieli czy na wzajemne oddziaływanie między nimi111. Przykłady te ilustrują potrzebę poprawy naszego rozumienia procesów ewolucyjnych i wdrożenia tej wiedzy w prognozy dotyczące wpływu, jaki wywierają zmiany klimatyczne.
Zakwaszenie oceanów konfrontuje mikroorganizmy morskie z poziomami PH wykraczającymi daleko poza ich niedawny historyczny zakres, co wpływa na ich homeostazę wewnątrzkomórkowego pH18,112. Gatunki, które są mniej biegłe w regulowaniu wewnętrznego pH odczuwają to bardziej, natomiast na zdolność regulacji wpływają czynniki takie jak wielkość organizmu, aktywność metaboliczna oraz tempo wzrostu112.
Niższe pH powoduje, że u bakterii i archeonów zmienia się ekspresja genów w sposób, który wspiera zachowanie komórek, nie zaś ich wzrost18. W mezokosmosach o niskiej biomasie, bakterie angażowały więcej środków do homeostazy pH niż bakterie w mezokosmosach bogatych w składniki pokarmowe o wysokiej biomamasie fitoplanktonu. W konsekwencji przewiduje się, że zakwaszenie oceanów zmieni sieć troficzną mikrobów poprzez zmiany skuteczności wzrostu komórek, obiegu węgla i ciągłe wahania energii, przy czym według prognoz największy wpływ będzie odczuwalny w regionach oligotroficznych, gdzie znajduje się najwięcej oceanów18. Eksperymentalne porównania wzrostu gatunku Synechococcus zarówno przy obecnych, jak i prognozowanych na przyszłość stężeniach pH wykazały wpływ nie tylko na sinice, ale także na wirusy znane jako cyjanofagi, które je infekują113.
Temperatura środowiska i szerokość geograficzna korelują z różnorodnością, rozmieszczeniem i/lub optymalną temperaturą (Topt) określonych morskich jednostek taksonomicznych, przy czym modele przewidują, że rosnące temperatury będą powodować przesuwanie się w kierunku bieguna społeczności przystosowanych do niskich temperatur52,114,115,116,117,118. Ustalono jednakże, że Topt fitoplanktonu z wód polarnych i wód strefy umiarkowanej jest znacząco wyższa niż temperatury środowiska, zaś model Eko-ewolucyjny przewidywał, że Topt dla fitoplanktonu w strefie tropikalnej będzie znacząco wyższa niż obserwowane wartości eksperymentalne116. Rozumienie jak dobrze mikroorganizmy przystosowane są do temperatury środowiska i prognozowanie w jaki sposób zareagują na ocieplenie wymagają oceny innych czynników niż Topt, która jest ogólnie rzecz biorąc słabym wskaźnikiem przystosowania fizjologicznego i ekologicznego mikroorganizmów ze środowisk o niskich temperaturach119.
Wiele czynników środowiskowych i fizjologicznych wpływa na reakcje i konkurencyjność mikroorganizmów w ich rodzimym środowisku. Na przykład podwyższone temperatury zwiększają syntezę białek w fitoplanktonie eukariotycznym jednocześnie zmniejszając stężenie rybosomów w komórce120. Jako że biomasa fitoplanktonu eukariotycznego wynosi ~1 Gt C (źródło13), a rybosomy są bogate w fosforan, zmiana ich stosunku azotu do fosforanu wywołana przez zmiany klimatu będzie miała wpływ na rozmieszczenie zasobów w oceanach na całym świecie120. Uważa się, że wzrost temperatury oceanów sprzyja bardziej rodzajom mniejszego planktonu niż większego, zmieniając przepływy biogeochemiczne takie jak eksport cząsteczek121. Przewiduje się, że podwyższone temperatury oceanów, zakwaszenie i zmniejszone zasoby środków pokarmowych zwiększą zewnątrzkomórkowe uwalnianie rozłożonej materii organicznej z fitoplanktonu, przy czym możliwe jest, że zmiany w pętli mikrobiologicznej spowodują większą produkcję mikrobiologiczną kosztem wyższych poziomów troficznych122. Ocieplenie może także złagodzić ograniczenie żelaza u sinic wiążących azot, a potencjalne konsekwencje dla nowego azotu dostarczanego do sieci troficznych przyszłych, cieplejszych oceanów mogą być poważne123. Baczną uwagę należy poświęcić kwestii wyliczenia ilościowego oraz interpretacji odpowiedzi mikroorganizmów środowiskowych na zmiany w ekosystemie i stresujące zjawiska powiązane ze zmianami klimatycznymi124,125. A zatem kluczowe pozostają pytania o skutki funkcjonalne przemieszczeń społeczności takie jak zmiany remineralizacji węgla w porównaniu z sekwestracją węgla, jak również obieg składników pokarmowych.
Biom lądowy
Biomasy naziemnej jest w przybliżeniu sto razy więcej niż biomasy morskiej, zaś rośliny lądowe stanowią dużą część całkowitej biomasy Ziemi15. Rośliny lądowe odpowiadają za mniej więcej połowę najważniejszej produkcji światowej netto30,67. Gleby zawierają ~2.000 miliardów ton węgla organicznego, czyli więcej niż łączne złoża żelaza w atmosferze i roślinności126. Całkowita liczba mikroorganizmów w środowiskach lądowych wynosi ~1029 i jest zbliżona do ich całkowitej liczby w środowiskach morskich8. Mikroorganizmy zasiedlające glebę regulują ilość węgla organicznego przechowywanego w glebie i uwalnianego z powrotem do atmosfery, a także wpływają pośrednio na magazynowanie węgla w roślinach i glebach poprzez dostarczanie makroelementów, które regulują produktywność (azot i fosfor)126,127. Rośliny dostarczają znaczną ilość węgla swoim symbiontom grzybom mikoryzowym, a w wielu ekosystemach grzyby mikoryzowe odpowiadają za przyswajanie znacznych ilości azotu i fosforu przez rośliny128.
Rośliny usuwają COz z atmosfery na drodze fotosyntezy i tworzą materię organiczną, która zasila ekosystemy lądowe. I odwrotnie, oddychanie autotroficzne przez rośliny (60 Pg C rocznie) i oddychanie heterotroficzne przez mikroorganizmy (60 Pg C rocznie) uwalniają CO2 z powrotem do atmosfery126,129. Temperatura wpływa na równowagę między tymi przeciwstawnymi procesami i tym samym na zdolność biosfery lądowej do wychwytywania i przechowywania wydzielanego węgla antropogenicznego (obecnie przechowywana jest w przybliżeniu jedna czwarta emitowanego węgla) (Ilustr. 1). Przewiduje się, że ocieplenie przyśpieszy uwalnianie węgla do atmosfery129.
Lasy zajmują ~30% powierzchni Ziemi, zawierają ~45% węgla lądowego, stanowią ~50% najważniejszej produkcji i sekwestrują do 25% antropogenicznego CO2 (źródła130,131). Łąki zajmują ~29% powierzchni lądowej132. Regiony niezalesione, suche i stepowe (47%) są ważne dla budżetu węgla i reagują na antropogeniczne zmiany klimatu inaczej niż tereny zalesione132,133. Jeziora stanowią ~4% terenów niezlodowaciałych134, zaś płytkie jeziora emitują znaczne ilości CH4 (źródła135,136). Torfowiska (zbutwiała ściółka leśna) zajmuje ~3% powierzchni Ziemi, przy czym ze względu na produktywność roślin wykraczającą poza butwienie nietknięte torfowiska funkcjonują jako globalny pochłaniacz dwutlenku węgla i zawierają ~30% światowego węgla w glebie137,138. W wiecznej zmarzlinie nagromadzenie węgla w materii organicznej (resztki roślin, zwierząt i mikroorganizmów) dalece przewyższa straty w oddychaniu tworząc największy lądowy pochłaniacz dwutlenku węgla139,140,141. Prognozuje się, że ocieplenie klimatu wynoszące 1,5-2°C (związane ze średnią światową temperaturą powierzchni w latach 1850-1900) zredukuje wieczną zmarzlinę o 28-53% (w porównaniu z poziomami w latach 1960-1990)142, to zaś sprawi, że duże rezerwuary węgla staną się dostępne do oddychania dla mikrobów oraz dla emisji gazów cieplarnianych.
Oceny górnych 10 cm gleby143 i profili całej gleby do głębokości 100 cm, która zawiera starsze zasoby węgla144, pokazują, że ocieplanie zwiększa utratę węgla do atmosfery. Wyjaśnienie różnicy w utracie węgla pomiędzy różnymi glebami wymagać będzie większego spektrum możliwych do przewidzenia zmiennych (poza zawartością materii organicznej w glebie, temperaturą, opadami, pH i zawartością gliny)145,146. Niemniej jednak prognozy wywodzące się z globalnych ocen reakcji na ocieplenie wskazują, że utrata węgla lądowego z powodu ocieplenia wywołuje sprzężenie zwrotne dodatnie, które przyśpiesza tempo zmian klimatycznych143, zwłaszcza w przypadku gleb zimnych i umiarkowanych przechowujących znaczną część światowego węgla w glebie147.
Mikroorganizmy wpływają na zmianę klimatu
Wyższe poziomy CO2 w atmosferze zwiększają najważniejszą produkcję i tym samym obszar ściółki leśnej oraz obumarłe korzenie148,149,150, co prowadzi do większej emisji węgla spowodowanej rozkładem mikrobiologicznym151. Wyższe temperatury sprzyjają szybszej dekompozycji lądowej materii organicznej152. Wpływ temperatury to nie tylko efekt kinetyczny przejawiający się w tempie reakcji drobnoustrojów, ale także skutki wkładu roślin stymulującego rozrost mikrobów152,153,154.
Kilka lokalnych czynników środowiskowych (takich jak skład społeczności drobnoustrojów, ciężar właściwy obumarłych drzew, dostępność tlenu i wilgotność) wpływa na wskaźniki aktywności mikrobiologicznej (na przykład na kolonizację drewna przez grzyby), co powoduje konieczność, aby przewidywania oparte o model systemu Ziemi i dotyczące utraty węgla w glebie na skutek ocieplenia klimatu uwzględniły lokalne metody regulacji procesów związanych z ekosystemem155. W tym kontekście dostępność roślinnych składników odżywczych wpływa na bilans netto węgla w lasach, przy czym lasy ubogie w składniki pokarmowe uwalniają więcej węgla niż lasy bogate w te składniki156. Respiracja mikrobów może być niższa w lasach bogatych w składniki odżywcze, jako że rośliny dostarczają mniej węgla (na przykład jako eksudaty) mikroorganizmom zasiedlającym strefę korzeniową157.
Rośliny uwalniają ~50% węgla stałego do gleby, która jest dostępna dla wzrostu drobnoustrojów158,159,160. Poza organizmami wykorzystującymi eksudaty jako źródło energii, wysięki mogą zakłócać asocjacje mineralno-organiczne wyzwalając związki organiczne z minerałów, które wykorzystywane są do respiracji mikrobów, przez co zwiększają uwalnianie węgla159. Istotność tego wzajemnego oddziaływania roślin i minerałów ilustruje znaczenie interakcji biotycznych i abiotycznych, jak również interakcji biotycznych (roślina-mikroorganizm), kiedy ocenia się wpływ zmian klimatu159. Modele termodynamiczne uwzględniające wzajemne oddziaływanie mikroorganizmów i wydzielanych enzymów z materią organiczną i minerałami stosowane są do prognozowania sprzężeń zwrotnych między zmianami klimatycznymi i węglem w glebie w odpowiedzi na rosnącą temperaturę; w jednym badaniu przewidziano bardziej zróżnicowane, ale słabsze sprzężenie zwrotne między klimatem i węglem w glebie na podstawie modelu termodynamicznego, nie zaś/niż na podstawie modeli statycznych160.
Dostępność materii organicznej w glebie dla degradacji mikrobiologicznej w przeciwieństwie do długotrwałego magazynowania zależy od wielu czynników środowiskowych, w tym właściwości mineralnych gleby, zakwaszenia i stanu redoks; dostępności wody, klimatu; i od rodzajów mikroorganizmów obecnych w glebie161. Właściwości materii organicznej, zwłaszcza złożoność substratu, wywierają wpływ na rozkład mikrobiologiczny. Ponadto, zdolność mikrobów do przedostawania się do materii organicznej różni się w zależności od typu gleby (różniących się na przykład zawartością gliny)162. Biorąc pod uwagę dostęp przewiduje się, że podwyższające się poziomy CO2 w atmosferze umożliwią rozkład mikrobiologiczny na szerszą skalę oraz mniejszą retencję węgla organicznego w glebie162.
Podwyższone stężenia CO2 zwiększają rywalizację roślin i mikroorganizmów o azot163. Zwierzęta roślinożerne (bezkręgowce i ssaki) mają wpływ na ilość materii organicznej, która powraca do gleby i tym samym na biomasę oraz aktywność mikrobiologiczną164. Dla przykładu koniki polne zmniejszają biomasę roślin i zapotrzebowanie roślin na azot jednocześnie zwiększając aktywność mikrobów163. Zmiany klimatu mogą zmniejszyć roślinożerność, a to może skutkować ogólnymi zmianami globalnego obiegu azotu i węgla, które zredukują lądową sekwestrację węgla163. Detrytofagi (na przykład dżdżownice) wpływają na emisję gazów cieplarnianych oddziałując pośrednio na rośliny (na przykład zwiększając żyzność gleby) i mikroorganizmy zasiedlające glebę165. Dżdżownice modyfikują gleby poprzez zasilanie, przekopywanie i odkładanie zbytecznych produktów przemiany materii. Beztlenowe środowisko jelita dżdżownic jest siedliskiem drobnoustrojów, które usuwają azot z azotanów i azotynów oraz produkują N2O.
Dżdżownice zwiększają żyzność gleby, a ich obecność może skutkować emisją gazów cieplarnianych netto165, chociaż łączne oddziaływanie podwyższonej temperatury i zmniejszonej ilości opadów na odżywianie detrytofagów i respirację mikrobów może redukować emisje166.
Odporna na próchnicę ściółka na torfowiskach (na przykład fenole przeciwdrobnoustrojowe i polisacharydy torfowców) hamuje rozkład mikrobiologiczny, zaś saturacja wody ogranicza wymianę tlenu oraz sprzyja wzrostowi anaerobów i uwalnianiu CO2 i CH4 (źródła137,167). Podwyższona temperatura i zmniejszona zawartość wody w glebie spowodowane zmianami klimatu oddziałują korzystnie na wzrost roślin naczyniowych (krzewów wrzosowatych), ale redukują produktywność mchów torfowych.
Zmiany w składzie ściółki roślin i związane z nimi procesy mikrobiologiczne (na przykład zmniejszone unieruchomienie azotu i zwiększona respiracja heterotroficzna) zmieniają torfowiska z pochłaniaczy dwutlenku węgla w jego źródła137. Erozja wiecznej zmarzliny na wybrzeżach doprowadzi do przemieszczenia się znacznych ilości węgla do oceanów, a przy tym dojdzie do potencjalnie wysokich emisji CO2 wskutek nasilonej remineralizacji mikrobiologicznej170, co z kolei wywoła sprzężenie zwrotne dodatnie, które przyśpieszy zmiany klimatyczne139,140,141,168,170,171. Topnienie wiecznej zmarzliny prowadzi do zwiększenia powierzchni gleb nawodnionych172, co sprzyja wytwarzaniu anaerobowego CH4 przez metanogeny i CO2 przez liczne mikroorganizmy. Produkcja jest powolna w porównaniu z metabolizmem w osuszonych glebach aerobowych, które zamiast CH4 uwalniają CO2. Jednakże trwające 7 lat badanie laboratoryjne dotyczące wytwarzania CO2 i CH4 wykazało, że kiedy społeczności metanogenów stały się aktywne w topniejącej wiecznej zmarzlinie, w warunkach beztlenowych powstały podobne ilości CO2 i CH4, przy czym przewidywano, że do końca stulecia emisje węgla ze środowisk anoksycznych doprowadzą do zmian klimatu na szerszą skalę niż emisje ze środowisk zawierających tlen172.
Prowadzone przez 15 lat badanie mezokosmosu symulujące środowiska jezior słodkowodnych wykazało, że skumulowane oddziaływanie eutrofizacji i ocieplenia może prowadzić do znacznego wzrostu wrzenia CH4 (kipienie pochodzące z nagromadzonego gazu)135. Ponieważ małe jeziora są podatne na eutrofizację i znajdują się przeważnie w regionach wrażliwych na warunki klimatyczne, należy ocenić na czym polega wkład mikroorganizmów jezior w światowe emisje gazów cieplarnianych135,136.
Zmiany klimatyczne wpływają na mikroorganizmy
Zmiany klimatu mogą wpływać na strukturę i zróżnicowanie społeczności mikrobiologicznych bezpośrednio (na przykład sezonowość i temperatura) lub pośrednio (na przykład skład roślinny, ściółka leśna i eksudaty). Różnorodność mikrobiologiczna gleb wpływa na zróżnicowanie roślinności i jest ważna dla funkcji spełnianych przez ekosystem, w tym dla obiegu węgla w przyrodzie173,174.
Zarówno krótkotrwałe ocieplenie w laboratorium, jak i długotrwałe (trwające ponad 50 lat) naturalne geotermalne ocieplenie początkowo zwiększały wzrost i respirację mikroorganizmów zasiedlających glebę prowadząc do uwalniania CO2 netto i późniejszego wyczerpania substratu oraz powodując zmniejszenie się biomasy i redukcję aktywności mikrobiologicznej175. Wskazuje to, że społeczności mikrobów nie przystosowują się łatwo do wyższych temperatur, zaś wynikający z tego wpływ na tempo reakcji i wyczerpanie substratu redukują ogólną utratę węgla175. W przeciwieństwie do tego, prowadzone przez 10 lat badanie wykazało, że społeczności zasiedlające glebę przystosowały się do rosnącej temperatury zmieniając skład i schematy wykorzystywania substratu, co prowadziło do mniejszej utraty węgla niż w przypadku braku adaptacji176. Znaczące zmiany w społecznościach bakterii i grzybów stwierdzono także w glebach leśnych z przekraczającym 20°C średnim zakresem rocznych temperatur177, jak również w odpowiedzi na ocieplanie w trwającym 9 lat badaniu gleb wysokotrawiastych prerii178.
W dwóch badaniach oceniono oddziaływanie podwyższonych temperatur na wskaźniki respiracji drobnoustrojów, a także na mechanizmy i rezultaty przystosowania179,180. W badaniach przeanalizowano szeroki zakres temperatur (-2 do 28°C), gleby suchego lądu (110 próbek) oraz gleby arktyczne, gleby strefy umiarkowanej i tropikalnej (22 próbki), jak również oceniono w jaki sposób społeczności reagują na trzy różne temperatury (~10-30°C). Adaptacja termiczna powiązana była z biofizycznymi właściwościami błon i enzymów komórkowych (odzwierciedlającymi kompromisy między aktywnością a stabilnością) oraz potencjałem genomowym mikroorganizmów (przy czym cieplejsze środowiska zasiedlają społeczności mikrobiologiczne o bardziej zróżnicowanym trybie życia179). Tempo respiracji na jednostkę biomasy było niższe w glebach w środowiskach o wyższej temperaturze, co wskazuje, że adaptacja termiczna może osłabić klimatyczne sprzężenie zwrotne dodatnie. Jednak, jako że oddychanie zależy od wielu powiązanych ze sobą czynników (nie tylko od jednej zmiennej takiej jak temperatura), takie mechaniczne rozumienie fizjologii mikrobów należy przedstawić na modelach biogeochemicznych możliwych klimatycznych sprzężeń zwrotnych dodatnich.
Jeśli chodzi o wzrost drobnoustrojów, ich reakcje na zmiany temperatury są skomplikowane i zróżnicowane181. Skuteczny wzrost mikrobów stanowi kryterium tego na ile efektywnie mikroorganizmy przetwarzają materię organiczną w biomasę, przy czym mniejsza efektywność oznacza, że większa ilość tlenu uwalniania jest do atmosfery182,183. Trwające tydzień badanie laboratoryjne ustaliło, że rosnąca temperatura prowadziła do zwiększenia wymiany mikrobiologicznej, ale nie do zmian w skuteczności wzrostu mikrobów, jak też prognozowało, że ocieplenie będzie sprzyjać gromadzeniu się węgla w glebie183. Badanie terenowe prowadzone na przestrzeni 18 lat wykazało, że efektywność drobnoustrojów była mniejsza przy wyższych temperaturach gleby, zaś rozkład opornego, skomplikowanego substratu wzrósł pod koniec tego okresu wraz z utratą netto węgla w glebie182.
Analogiczne w trwającym 26 lat badaniu, które dotyczyło ocieplenia gleb leśnych, w rozkładzie materii organicznej i uwalnianiu CO2 nastąpiło krótkotrwałe odchylenie, prowadzące do zmian w składzie społeczności mikrobiologicznej i w efektywności wykorzystywania węgla, zredukowanej biomasy mikrobiologicznej i zmniejszonej ilości węgla dostępnego dla mikrobów184. Ogólnie rzecz biorąc badanie to prognozowało, że antropogeniczne zmiany klimatu spowodują długoterminowe, rosnące i nieprzerwane uwalnianie węgla184. Podobne przewidywania wynikają z modeli systemowych Ziemi, które imitują reakcje fizjologiczne drobnoustrojów185 lub uwzględniają wpływ zamarzania i topnienia gleb w zimnym klimacie186.
Zmiany klimatyczne bezpośrednio i pośrednio wpływają na społeczności mikrobiologiczne i na ich funkcje poprzez kilka powiązanych ze sobą czynników, takich jak temperatura, opady, właściwości gleby oraz wkład roślin. Jako że mikroorganizmy zasiedlające glebę na pustyniach zawierają ograniczoną ilość węgla, rosnący dopływ węgla z roślin sprzyja transformacji związków azotowych, biomasie mikrobiologicznej, różnorodności (na przykład grzybów), funkcjonowaniu enzymów oraz wykorzystaniu opornej materii organicznej133. Chociaż zmiany te mogą usprawniać respirację i utratę netto węgla z gleby, konkretne właściwości obszarów suchych i stepowych mogą oznaczać, że możliwe byłoby ich funkcjonowanie jako pochłaniaczy węgla133. Jednak badanie 19 obszarów trawiastych strefy umiarkowanej wykazało, że sezonowe różnice w opadach deszczu ograniczają gromadzenie się biomasy. Aby lepiej zrozumieć naziemne reakcje roślina-biomasa na poziomy CO2 i na sezonowe opady, potrzebna nam także większa wiedza na temat reakcji i funkcji społeczności mikrobiologicznych.
Dane dotyczące metagenomów, w tym genomów poskładanych z metagenomów, dostarczają wiedzy o kluczowych grupach drobnoustrojów, które metabolizują materię organiczną i uwalniają CO2 i CH4 oraz wiążą te grupy z biogeochemią topniejącej wiecznej zmarzliny187,188,189,190,191. Społeczności mikrobiologiczne zasiedlające tundrę zmieniają się w warstwie gleby wiecznej zmarzliny w następstwie ocieplenia192. W ciągu półtora roku od ocieplenia potencjał funkcjonalny społeczności mikrobiologicznych znacząco się zmienił, przy czym coraz większa liczba genów zaangażowana była w aerobowy i anaerobowy rozkład oraz obieg składników odżywczych w przyrodzie. Chociaż metabolizm mikrobiologiczny stymuluje najważniejszą produktywność roślin, równowaga między respiracją mikrobów a najważniejszą produktywnością skutkuje uwalnianiem netto węgla do atmosfery192. Kiedy lasy rozprzestrzeniają się na ocieplające się obszary tundry, wzrost roślin może powodować utratę netto węgla, być może na skutek stymulowania przez eksudaty rozkładu mikrobiologicznego węgla w macierzystej glebie153,193. Mimo że istnieją sprawozdania mówiące o kumulowaniu się węgla w wyniku ocieplenia (na przykład źródło183), większość badań opisuje reakcje społeczności mikrobiologicznych, które skutkują utratą węgla.
Gwałtowne ocieplenie Półwyspu Antarktycznego i powiązanych z nim wysp przyniosło skutek w postaci rozszerzenia obszaru występowania śmiałka antarktycznego (Deschampsia antarctica), jako że wypiera on inne gatunki występujące naturalnie na danym obszarze (na przykład mech Sanionia uncinata) dzięki wyjątkowej zdolności swoich korzeni do pozyskiwania peptyd i tym samym azotu194. Zdolność traw do konkurowania zależy od trawienia przez drobnoustroje białek pozakomórkowych oraz wytwarzania aminokwasów, azotanu i amoniaku194. Ponieważ cieplejsze gleby na tym obszarze są siedliskiem większej różnorodności grzybów, przewiduje się, że zmiany klimatu spowodują zmiany w społecznościach grzybów, które będą miały wpływ na obieg składników odżywczych w przyrodzie i na najważniejszą produktywność195. Zróżnicowanie sinic i produkcja toksyn na matach bentonicznych zarówno na Półwyspie Antarktycznym, jak i w Arktyce wzrosły w ciągu ostatnich 6 miesięcy kontaktu z szybko rosnącymi temperaturami196. Zmiana na gatunki wytwarzające toksyny lub rosnąca produkcja toksyn przez istniejące gatunki mogłyby zaszkodzić polarnym jeziorom słodkowodnym, gdzie sinice są często dominującym bentonicznym gatunkiem wytwarzającym.
Prawdopodobne jest, że zmiany klimatyczne zwiększą częstotliwość, nasilenie i czas trwania zakwitu sinic w wielu jeziorach eutroficznych, sztucznych zbiornikach wodnych i w ujściach rzek do morza197,198. Sinice tworzące zakwity produkują różnorodne neurotoksyny, hepatotoksyny i dermatoksyny, które mogą być zgubne dla ptaków i ssaków (w tym dla ptactwa wodnego, bydła i psów) i mogą zagrozić wykorzystywaniu wód w celach rekreacyjnych, produkcji wody pitnej, irygacji rolniczej oraz rybołówstwu198. Toksyczne sinice powodują poważne problemy z jakością wody na przykład na Jeziorze Tai Hu (Chiny), na Jeziorze Erie (USA), na Jeziorze Okeechobee (USA), na Jeziorze Wiktorii (Afryka) oraz na Morzu Bałtyckim198,199,200.
Zmiany klimatu sprzyjają zakwitom sinic zarówno bezpośrednio, jak i pośrednio198. Wiele sinic tworzących zakwity może rozwijać się przy stosunkowo wysokich temperaturach201. Rosnąca stratyfikacja termiczna jezior i sztucznych zbiorników wodnych umożliwia prężnym sinicom utrzymywanie się na powierzchni i tworzenie gęstych zakwitów powierzchniowych, co daje im lepszy dostęp do światła i dzięki temu przewagę selektywną nad nieprężnymi organizmami fitoplanktonowymi202,203. Przewlekłe susze w okresie letnim wydłużają czas retencji wody w sztucznych zbiornikach, rzekach i ujściach rzek do morza, zaś te stojące ciepłe wody mogą zapewniać idealne warunki do rozwoju zakwitu sinic204.
Zdolność szkodliwego rodzaju sinic Microcystis do adaptacji do podwyższonych poziomów CO2 wykazano zarówno w doświadczeniach laboratoryjnych, jak i w terenie205. Microcystis spp. absorbują CO2 i HCO3 oraz kumulują węgiel nieorganiczny w karboksysomach, przy czym ustalono, że konkurencja między szczepami zależy od stężenia węgla nieorganicznego. W rezultacie oczekuje się, że podwyższone poziomy CO2 będą miały wpływ na skład szczepów zakwitów sinic205.
Rolnictwo
Według Banku Światowego (dane Banku Światowego dotyczące ziemi uprawnej), niemal 40% środowiska lądowego wykorzystywane jest do produkcji rolnej. Prognozuje się, że stosunek ten będzie wzrastać prowadząc do istotnych zmian w obiegu składników odżywczych w glebie, m.in. węgla, azotu i fosforu. Ponadto zmiany te wiążą się ze znaczącą utratą bioróżnorodności206, w tym mikroorganizmów207. Coraz większym zainteresowaniem cieszy się wykorzystywanie mikroorganizmów zasocjowanych z roślinami lub zwierzętami do wzmocnienia zrównoważonego rozwoju rolnictwa oraz do zminimalizowania skutków zmian klimatycznych dla produkcji żywności, jednak praktykowanie tego wymaga lepszego zrozumienia sposobu w jaki zmiany klimatu będą wpływać na mikroorganizmy.
Mikroorganizmy wpływają na zmiany klimatu
Oprócz antropogenicznej produkcji metanu związanej z paliwami kopalnymi208, metanogeny wytwarzają metan w naturalnych i sztucznych środowiskach anaerobowych (osady, grunty nawodnione takie jak pola ryżowe, przewód pokarmowy zwierząt (zwłaszcza przeżuwaczy), urządzenia wodociągowo-kanalizacyjne i oczyszczalnie biogazowe)(Ilustracja 2). Głównymi pochłaniaczami CH4 są utlenienie atmosferyczne i utlenienie mikrobiologiczne w glebach, osadach i w wodzie208. Poziomy CH4 w atmosferze wzrosły gwałtownie w ostatnich latach (2014-2017), ale powody tego stanu rzeczy są jak dotąd niejasne, choć wiążą się z coraz większymi emisjami związanymi z przemysłem metanogenowym i/lub paliwami kopalnymi i/lub ze zredukowanym utlenieniem CH4 atmosferycznego, tym samym stanowiąc istotne zagrożenie dla kontroli ocieplania się klimatu209.
Ilustracja 2: Rolnictwo i inne rodzaje działalności człowieka mające wpływ na mikroorganizmy.
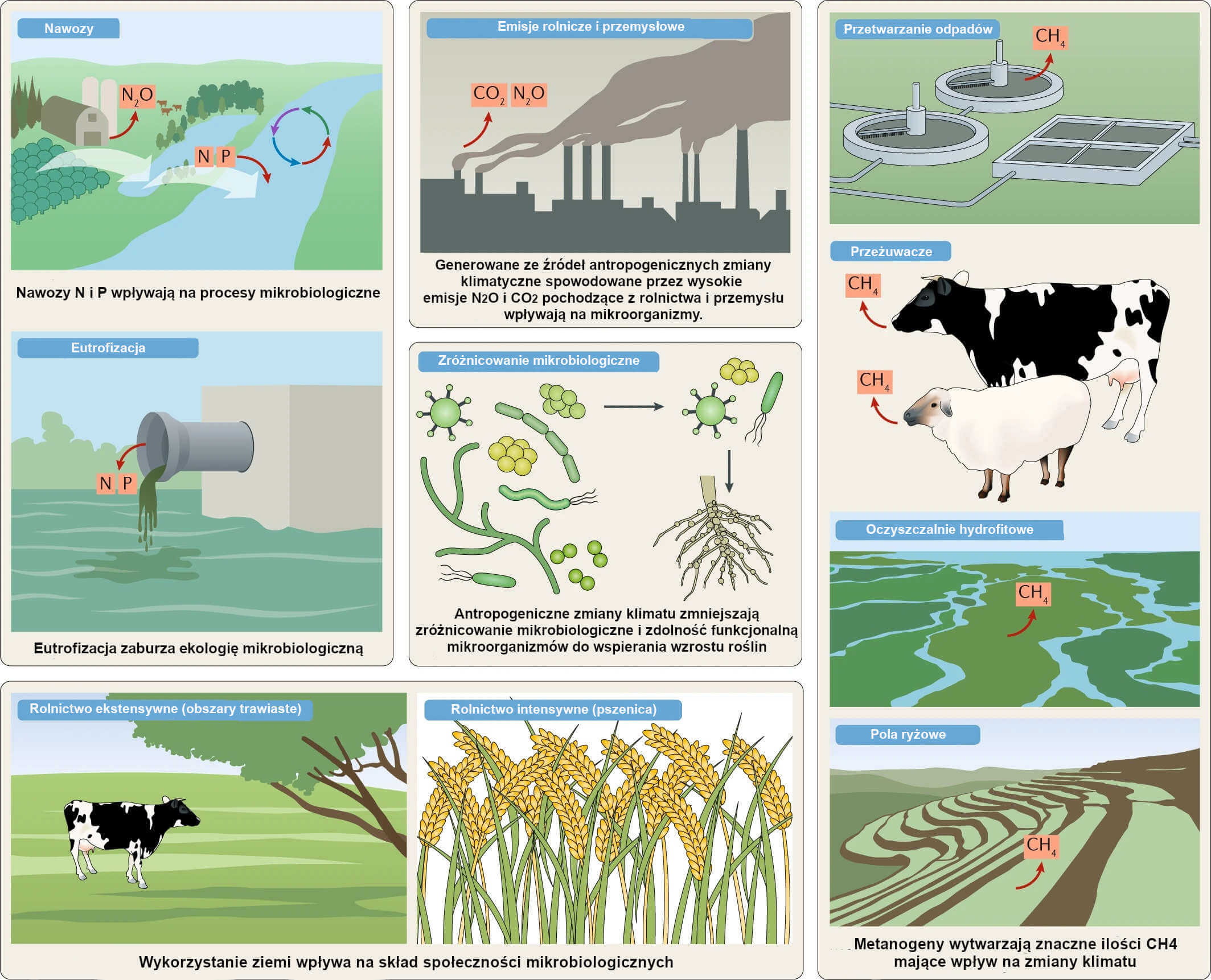
Działalność rolnicza wpływa na społeczności mikrobiologiczne w określony sposób. Wykorzystanie ziemi (na przykład typ roślin) i źródła zanieczyszczenia (na przykład nawozy) zakłócają skład i funkcjonowanie społeczności mikrobiologicznych, tym samym wpływając na transformacje naturalnego obiegu węgla, azotu i fosforu w przyrodzie. Metanogeny produkują znaczne ilości metanu bezpośrednio z wykorzystaniem zwierząt przeżuwających (na przykład bydła, owiec i kóz) i nawodnionych gleb o warunkach anaerobowych (na przykład pola ryżowe i oczyszczalnie hydrofitowe). Działalność człowieka, która powoduje redukcję zróżnicowania mikrobiologicznego zmniejsza także zdolność drobnoustrojów do wspierania wzrostu roślin.
Połowa populacji świata żywi się ryżem210, a pola ryżowe odpowiadają za ~20% emisji CH4 pochodzących z rolnictwa, mimo że zajmują zaledwie ~10% gruntów rolnych. Przewiduje się, że do końca tego stulecia antropogeniczne zmiany klimatu zwiększą dwukrotnie emisje CH4 pochodzące z upraw ryżu210. Zwierzęta przeżuwające są największym pojedynczym źródłem antropogenicznych emisji CH4, przy czym ślad węglowy produkcji mięsa przeżuwaczy jest 19-48 razy większy niż w przypadku bogatej w białko żywności opartej o rośliny211. Nawet produkcja mięsa zwierząt nieprzeżuwających (takich jak świnie, drób i ryby) wytwarza 3-10 razy więcej CH4 niż żywności pochodzenia roślinnego o wysokiej zawartości białka211.
Spalanie paliw kopalnych oraz stosowanie nawozów zdecydowanie zwiększyły występowanie azotu w atmosferze zaburzając globalne procesy biogeochemiczne i zagrażając stabilności ekosystemu212,213. Rolnictwo jest największym emitentem silnie działającego gazu cieplarnianego N2O, który uwalnia się na drodze oksydacji mikrobiologicznej i redukcji azotu214. Enzym reduktaza N2O w bakteriach zasiedlających strefę korzeniową (w brodawkach korzeniowych) i inne mikroorganizmy w glebie mogą także przetwarzać N2O w N2 (nie w gaz cieplarniany). Zmiany klimatu zakłócają tempo w jakim zachodzą mikrobiologiczne transformacje azotu (rozkład, mineralizacja, nitryfikacja, denitryfikacja i wiązanie) i w jakim wydalają N2O (źródła213). Istnieje pilna potrzeba poznania wpływu zmian klimatu i innych przejawów działalności człowieka na mikrobiologiczne transformacje związków azotu.
Zmiany klimatu wpływają na mikroorganizmy
Uprawa roślin waha się od uprawy prowadzonej zgodnie z zasadami rolnictwa ekstensywnego (niewielki wkład pracy, nawozów i kapitału) do prowadzonej zgodnie z zasadami rolnictwa intensywnego (duży wkład). Rosnąca temperatura i susza wpływają silnie na możliwość uprawy roślin215. Oparte na grzybach sieci troficzne w glebie występują powszechnie w rolnictwie ekstensywnym (na przykład pastwiska) i są bardziej zdolne do przystosowania się do suszy niż sieci troficzne oparte na bakteriach, które są częste w systemach intensywnych (na przykład pszenica)216,217. Światowa ocena górnej warstwy gleby wykazała, że grzyby i bakterie w glebie zajmują określone miejsca i reagują różnie na opady i pH gleby wskazując, że zmiany klimatu miałyby zróżnicowany wpływ na ich liczebność, różnorodność i funkcje218. Jałowość, która według prognoz ma się zwiększyć na skutek zmian klimatu, redukuje różnorodność i liczebność bakterii oraz grzybów na suchych terenach całego świata219. Redukcja różnorodności mikrobiologicznej w glebie zmniejsza ogólny potencjał funkcjonalny społeczności drobnoustrojów ograniczając tym samym ich zdolność do wspierania wzrostu roślin173.
Łączne oddziaływanie zmian klimatu i eutrofizacji w wyniku stosowania nawozów może mieć istotny, potencjalnie niemożliwy do przewidzenia wpływ na konkurencyjność mikroorganizmów. Dla przykładu nadmiar substancji biogennych zazwyczaj sprzyja szkodliwym zakwitom alg, ale inny skutek zaobserwowano we względnie głębokim Jeziorze Zuryskim220. Redukcja dopływu fosforu z nawozów zmniejszyła zakwity fitoplanktonu eukariotycznego, lecz doprowadziła do wzrostu wskaźnika azotu do fosforu i w ten sposób dominować zaczęły nie wiążące azotu sinice Planktothrix rubescens220. Jeżeli nie występują skuteczne drapieżniki, roczne mieszanie odgrywa ważną rolę w kontrolowaniu populacji sinic. Jednak ocieplenie nasiliło stratyfikację cieplną i zredukowało mieszanie, a to z kolei spowodowało zwiększenie się zdolności do przetrwania toksycznych sinic220.
Choroby zakaźne
Zmiany klimatu mają wpływ na występowanie i rozprzestrzenianie się chorób wśród morskiej i lądowej flory i fauny221 (Ilustracja 3) w zależności od różnorodnych aspektów socjoekonomicznych i środowiskowych oraz czynników szczególnych dla żywiciela i patogenu222. Zrozumienie mechanizmów rozprzestrzeniania się choroby i opracowywanie skutecznych strategii kontroli wymaga znajomości ekologii patogenów, ich wektorów i ich żywicieli, a także wpływu czynników środowiskowych i związanych z rozprzestrzenianiem się223 (Tabela 1). Przykładowo występuje silny związek między rosnącymi temperaturami powierzchni morza a chorobami koralowców i mimo że mechanizmy choroby nie są absolutnie jasne dla wszystkich różniących się od siebie objawów, powiązania z patogenami mikroorganizmów istnieją224,225,226. Wartości szczytowe zachorowalności zbiegają się z cyklicznością Oscylacji Południowej El Niño (ang. El Niño Southern Oscillation, ENSO)227. W szczególności u niektórych gatunków koralowców ocieplanie się oceanu może zmienić ich mikrobiom zaburzając równowagę żywiciel-symbiont i wprowadzając zmiany w mechanizmach obronnych oraz ścieżkach obiegu składników odżywczych, które mogą przyczyniać się do bielenia i choroby99. Zakwaszenie oceanów może również bezpośrednio powodować uszkodzenie tkanek u organizmów takich jak ryby, potencjalnie przyczyniając się do osłabienia układu odpornościowego, które stwarza warunki do inwazji bakterii228.
Ilustracja 3. Zmiany klimatu nasilają wpływ patogenów.
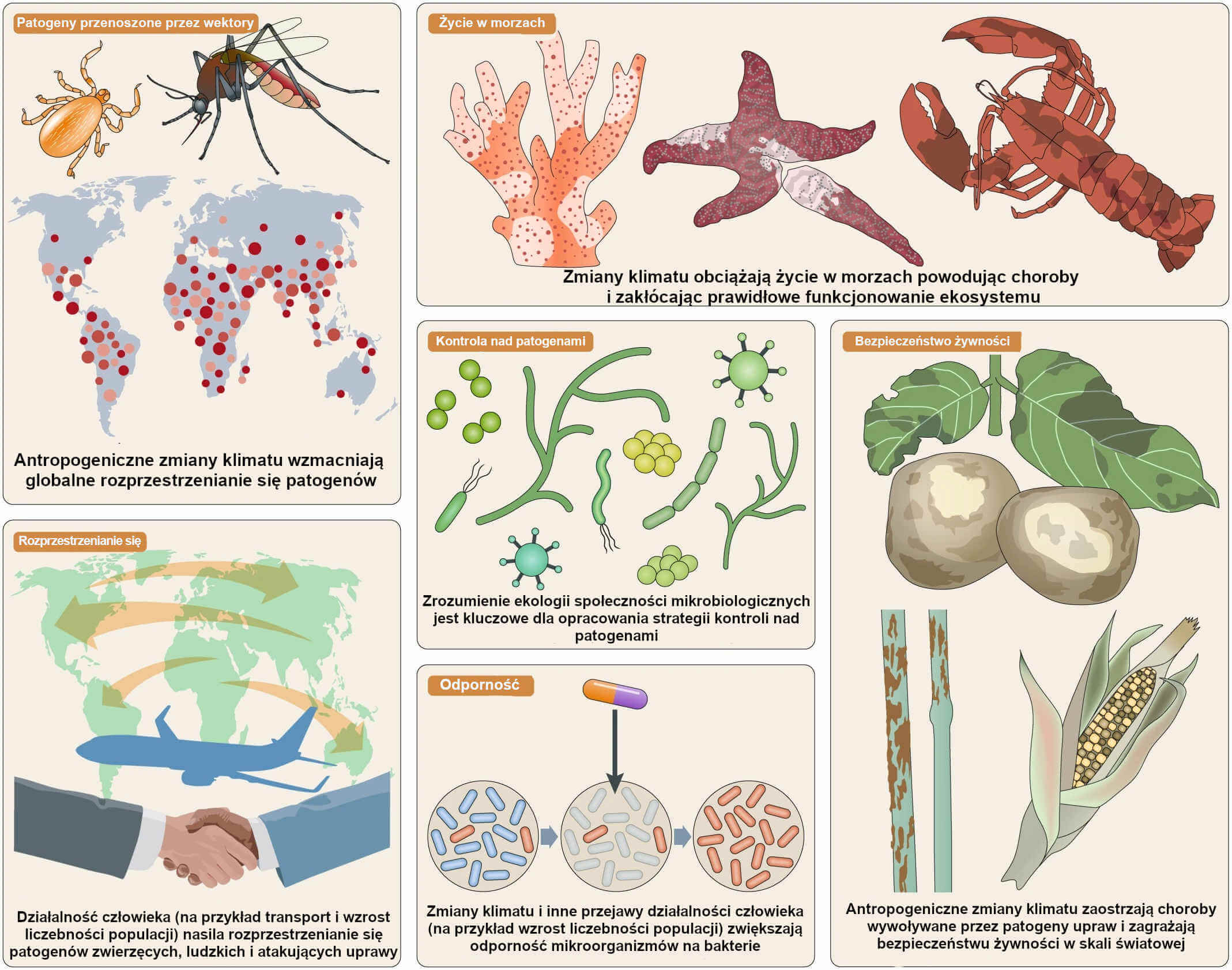
Antropogeniczne zmiany klimatu obciążają występujące naturalnie formy życia zwiększając tym samym zdolność patogenów do wywoływania chorób na coraz większą skalę. Wpływ na akwakulturę, zwierzęta hodowlane i uprawy zagraża zaopatrzeniu całego świata w żywność. Działalność człowieka jak np. wzrost liczebności populacji i transport, w połączeniu ze zmianami klimatu, zwiększa odporność patogenów na antybiotyki oraz rozprzestrzenianie się czynników chorobotwórczych pochodzenia wodnego i patogenów przenoszonych prze wektory, przez co ludzie, inne zwierzęta i rośliny częściej zapadają na choroby.
Tabela 1 Wpływ czynników klimatycznych i środowiskowych na przenoszenie patogenów
Zobacz tabelę w pełnym rozmiarze
Populacja gwiazdy morskiej wzdłuż liczącej ~3000 km części północnoamerykańskiego zachodniego wybrzeża zmniejszyła się o 80-100%, przy czym szczytowe spadki wystąpiły podczas anormalnych wzrostów temperatur powierzchni oceanu229. Jako że gwiazdy morskie są ważnymi drapieżnikami polującymi na jeżowce, ich brak może powodować kaskadę troficzną, która oddziałuje na lasy wodorostów i powiązaną bioróżnorodność morską229,230. Uwzględniając wpływ ocieplania się oceanów na aktywność patogenów opracowano systemy monitorowania temperatury dla szerokiej gamy organizmów morskich, w tym koralowców, gąbek, ostryg, homarów i innych skorupiaków, gwiazd morskich, ryb oraz traw morskich231.
Patogeny mogą nasilać wymieranie drzewostanu spowodowane suszą i stresem cieplnym232. Kiedy rozważa się reakcję roślin uprawnych na patogeny, w tym na ilości CO2, zmiany klimatyczne, stan roślin oraz interakcje roślina-patogeny konkretne dla danego gatunku, istotna jest różnorodność współdziających ze sobą czynników233. Znaczna liczba mikroorganizmów powoduje choroby roślin (grzyby, bakterie, wirusy, wiroidy i lęgniowce) i z tej przyczyny mogą one wpływać na produkcję roślin uprawnych, wywoływać klęski głodu (na przykład lęgniowiec Phytophthora infestans spowodował ziemniaczaną klęskę głodu w Irlandii) i zagrażać bezpieczeństwu żywności233. Ocena ponad 600 szkodników (insektów i nicieni) i patogenów roślin uprawnych od 1960 roku wykazała ekspansję w kierunku biegunów, którą przypisuje się zmianom klimatu233. Transport i pierwszy kontakt z różnymi gatunkami powodują, że patogeny łatwiej się rozprzestrzeniają i pojawiają się choroby, na co wpływa też oddziaływanie pogody na rozsiewanie się i warunki środowiskowe do wzrostu233.
Zmiany klimatyczne mogą zwiększać ryzyko chorób zmieniając przystosowanie się żywiciela i pasożyta234. W przypadku zwierząt zmiennocieplnych (jak na przykład płazy) temperatura może zwiększać podatność na infekcje prawdopodobnie w wyniku zaburzenia odpowiedzi immunologicznych234,235. Miesięczne i dzienne niemożliwe do przewidzenia wahania temperatury środowiska nasilają podatność kubańskiej żaby drzewnej na patogenny grzyb z gromady skoczkowców Batrachochytrium dendrobatidis. Wpływ rosnącej temperatury na infekcje kontrastuje ze zmniejszającą się zdolnością do wzrostu tego grzyba w czystej kulturze odzwierciedlając wagę oceny reakcji żywiciel-patogen (zamiast wnioskować na podstawie badań tempa wzrostu wyizolowanych drobnoustrojów) przy wydawaniu opinii na temat znaczenia zmian klimatu234.
Prognozuje się, że zmiany klimatyczne zwiększą wskaźniki odporności na antybiotyki niektórych ludzkich patogenów236. Dane z lat 2013-2015 wskazują, że wzrost dziennej temperatury minimalnej o 10°C (co jest możliwe w niektórych częściach USA przed końcem obecnego stulecia) doprowadzi do zwiększenia się wskaźników odporności na antybiotyki Escherichia coli, Klebsiella pneumoniae i Staphylococcus aureus o 2-4% (w przypadku niektórych antybiotyków do 10%)236. Potencjalne główne mechanizmy obejmują podwyższone temperatury ułatwiające horyzontalny transfer genów ruchomych genetycznych elemenów odporności oraz zwiększone tempo wzrostu patogenów wspierające zdolność do przetrwania w danym środowisku, transport i przenoszenie236. Wzrost liczebności populacji, który potęguje zmiany klimatu również stanowi istotny czynnik przyczyniający się do rozwoju odporności236.
Patogeny wektorowe, patogeny przenoszone drogą pokarmową, patogeny przenoszone drogą powietrzną i przenoszone przez wodę oraz inne patogeny obecne w środowisku mogą być szczególnie podatne na skutki zmian klimatycznych237,238,239,240 (Tabela 1). W przypadku chorób przenoszonych przez wektory zmiany klimatu wpływają na rozmieszczenie wektorów i stąd na skalę przenoszenia chorób, a także na skuteczność, z jaką wektory przenoszą patogeny. Skuteczność zależy od czasu pomiędzy żywieniem się wektora na zainfekowanym żywicielu a chwilą, w której wektor sam staje się zakaźny. Przy wyższych temperaturach czas ten może być znacząco zredukowany, co zapewnia więcej okazji do przenoszenia w okresie życia wektora. Niektóre schorzenia przenoszone przez wektory – takie jak choroba niebieskiego języka, ważna pod względem ekonomicznym choroba wirusowa trzody – pojawiły się już w Europie w odpowiedzi na zmiany klimatu, przy czym przewiduje się, że w przyszłości wybuchy epidemii tej choroby będą częstsze i zakrojone na szerszą skalę241. W przypadku niektórych infekcji przenoszonych drogą wodną przez patogenne Vibrio spp. rozprzestrzenianie się w kierunku biegunów koreluje z rosnącą globalną temperaturą i mniejszym zasoleniem środowisk wodnych w rejonach wybrzeża (takich jak ujścia rzek) spowodowanymi coraz większą ilością opadów242. Te zmienione warunki mogą sprzyjać wzrostowi Vibrio spp. w danym środowisku242. Rosnące temperatury powierzchni morza są także związane z coraz częstszymi przypadkami zakażenia Vibrio cholerae w Bangladeszu243, infekcjami wywołanymi przez kilka patogennych dla człowieka Vibrio spp. w rejonie Morza Bałtyckiego242 i bogactwem Vibrio spp. (w tym ludzkich patogenów) na Północnym Atlantyku i na Morzu Północnym244.
Malaria i denga to dwie choroby przenoszone przez wektory, co do których wiadomo, że są wysoce wrażliwe na warunki klimatyczne i z tego powodu przewiduje się, że ich rozmieszczenie przestrzenne będzie się zmieniać w odpowiedzi na zmiany klimatu4,141,245. Zmiany te mogą ułatwiać rozprzestrzenianie się patogenów przenoszonych przez wektory wydłużając okres transmisji, przyspieszając tempo rozmnażania się patogenów w wektorze oraz zwiększając wzrost liczebności i obszar występowania komarów. Dotyczy to zwłaszcza Aedes aegypti, najważniejszego wektora wirusów dengi, ziki, czikungunii i żółtej febry, którego występowanie obecnie ogranicza się do regionów tropikalnych i subtropikalnych, ponieważ nie jest w stanie przetrwać mroźnych zim. Przewiduje się, że w połączeniu z innymi chorobami przenoszonymi przez komary (jak np. gorączka zachodniego Nilu czy japońskie zapalenie mózgu) i chorobami przenoszonymi przez kleszcze (jak borielioza z Lyme) miliony ludzi jest od niedawna narażonych na ryzyko z powodu zmian klimatycznych4,238,246,247,248,249.
Na wiele chorób zakaźnych, w tym kilka schorzeń przenoszonych przez wektory i pochodzenia wodnego, silny wpływ wywiera zmienność klimatu spowodowana przez zjawiska klimatyczne na dużą skalę, takie jak ENSO, która zakłóca typowe schematy opadów deszczu i zmienia temperatury w mniej więcej jednej trzeciej świata co kilka lat. Powiązania z ENSO zgłasza się w przypadku malarii, dengi, choroby wywołanej wirusem Zika, cholery, dżumy, afrykańskiego pomoru koni i wielu innych ważnych chorób ludzkich oraz zwierzęcych250,251,252,253,254.
Adaptacja mikroorganizmów do ich lokalnego środowiska stanowi przedmiot badań w mniejszym stopniu niż w odniesieniu do zwierząt (w tym ludzi) i roślin, choć bada się mechanizmy i konsekwencje przystosowania się w naturalnych i eksperymentalnych populacjach mikrobiologicznych255. Patogeny wirusowe, bakteryjne i grzybowe roślin i zwierząt (w tym roślin uprawnych, ludzi i żywego inwentarza) przystosowują się do czynników abiotycznych i biotycznych (takich jak temperatura, pestycydy, interakcje między mikroorganizmami i odporność żywiciela) w sposób, który wpływa na funkcjonowanie ekosystemu, ludzkie zdrowie i bezpieczeństwo żywności255. Cykliczne sprzężenie zwrotne reakcji mikrobów z działalnością człowieka dobrze ilustrują modele adaptacji patogennych grzybów rolniczych256. Ponieważ ekosystemy rolnicze posiadają wspólne cechy na całym świecie (na przykład irygacja, stosowanie nawozów i rośliny uprawne), a przemieszczanie się ludzi i transport materiału roślinnego z łatwością doprowadzają do rozprzestrzeniania się patogenów roślin uprawnych, patogeny przystosowane do warunków rolniczych mają większy potencjał wywoływania epidemii i stanowią większe zagrożenie dla produkcji roślin uprawnych niż szczepy występująe naturalnie256. Zdolność patogenów grzybowych do ekspansji i opanowywania nowych środowisk uzyskiwana na drodze ewolucji tak, aby mogły tolerować wyższe temperatury zwiększa zagrożenie, jakie patogeny grzybowe stanowią zarówno dla ekosystemu naturalnego, jak i rolniczego257.
Łagodzenie zmian klimatycznych przez mikroorganizmy
Lepsze zrozumienie interakcji mikrobiologicznych pomogłoby w opracowaniu środków zapobiegawczych służących łagodzeniu i kontroli zmian klimatu oraz ich skutków (patrz także źródło7). Dla przykładu zrozumienie w jaki sposób komary reagują na bakterię Wolbachia (powszechny symbiont stawonogów) przyniosło skutek w postaci spadku transmisji wirusów ziki, dengi i czikungunii poprzez wprowadzenie Wolbachia do populacji komarów A. aegypti i wypuszczeniu tych komarów do środowiska258. W rolnictwie coraz lepsze rozumienie ekofizjologii mikroorganizmów, które redukują N2O do nieszkodliwego N2 daje możliwości zmniejszenia emisji214,259. Wykorzystanie szczepów bakterii o wyższej aktywności reduktazy N2O obniżyło emisje N2O z soi, przy czym zarówno naturalne, jak i zmodyfikowane genetycznie szczepy z wyższą aktywnością reduktazy N2O zapewniają drogi minimalizowania emisji N2O214. Manipulowanie mikrobiomem żwacza260 i programy hodowlane, które celują w czynniki genetyczne żywiciela zmieniające reakcje społeczności mikrobiologicznej262 to niektóre ze sposobów redukcji emisji metanu pochodzącego od bydła. W tym drugim przypadku celem byłaby produkcja linii bydła, które podtrzymują społeczności mikrobiologiczne wytwarzające mniej metanu bez wpływu na zdrowie i produktywność zwierząt261. Białka grzybów mogą zastąpić mięso zmniejszając ślad węglowy diety262.
Biowęgiel stanowi przykład rozwiązania z dziedziny rolnictwa służący szeroko zakrojonemu i pośredniemu łagodzeniu wpływu zmian klimatu na mikroorganizmy. Biowęgiel produkuje się na drodze termochemicznego przetwarzania biomasy w warunkach ograniczonego dostępu tlenu i poprawia on stabilizację i gromadzenie się materii organicznej w glebach bogatych w żelazo263. Biowęgiel wzmacnia retencję materii organicznej zmniejszając mineralizację mikrobiologiczną i redukując wpływ wysięków z korzeni na uwalnianie materiału organicznego z minerałów tym samym wspierając wzrost traw i minimalizując uwalniania się węgla263.
Podejściem do minimalizowania tych problemów potencjalnie zakrojonym na szeroką skalę jest wykorzystywanie oczyszczalni hydrofitowych do produkcji biopaliwa celulozowego z zastosowaniem nieużytecznego azotu pochodzącego z oczyszczania ścieków; gdyby wykorzystywano wszystkie odpady w Chinach, mogłoby to zapewnić ekwiwalent 7% zużycia benzyny w tym kraju264. Tak poważne zmiany w wykorzystywaniu oczyszczalni hydrofitowych wymagałyby opisu i optymalizacji ich zasadniczych konsorcjów mikrobiologicznych służących zarządzaniu emitowanymi przez nie gazami cieplarnianymi i zwiększaniu korzyści dla środowiska265.
Biotechnologia mikrobiologiczna może być źródłem rozwiązań dla zrównoważonego rozwoju266 między innymi w zakresie zapewniania (na przykład żywności) i regulowania (na przykład chorób lub emisji i wychwytywania gazów cieplarnianych) usług ekosystemowych dla ludzi, zwierząt i roślin. Technologie mikrobiologiczne dostarczają rozwiązań praktycznych (substancje chemiczne, materiały, energię i rekultywację) służących osiągnięciu wielu spośród 17 Celów Zrównoważonego Rozwoju Organizacji Narodów Zjednoczonych dotyczących ubóstwa, głodu, zdrowia, czystej wody, czystej energii, rozwoju gospodarczego, innowacji przemysłowych, miast zrównoważonych, odpowiedzialnej konsumpcji, działań w zakresie klimatu, życia pod wodą i życia na lądzie6 (Ramka 1). Zwiększanie poparcia dla takich działań bez wątpienia ułatwi poprawa powszechnego rozumienia kluczowych ról jakie mikroorganizmy odgrywają w globalnym ociepleniu, które osiągnięto by na drodze uzyskania elokwencji mikrobiologicznej przez społeczeństwo7.
Wnioski
Drobnoustroje mają znaczący wkład w sekwestrację węgla, zwłaszcza morskiego fitoplanktonu, który wiąże taką samą ilość CO2 netto, co rośliny lądowe. Z tej przyczyny zmiany w środowisku, które wpływają na morską fotosyntezę mikrobiologiczną i na późniejsze przechowywanie stałych węglowych w głębokich wodach mają kardynalne znaczenie dla globalnego obiegu węgla w przyrodzie. Mikroby przyczyniają się również znacząco do emisji gazów cieplarnianych poprzez respirację heterotroficzną (CO2), metanogenezę (CH4) i usuwania azotu z azotanów i azotynów (N2O).
Wiele czynników ma wpływ na równowagę między wychwytywaniem mikrobiologicznych gazów cieplarnianych a emisją, m.in. biom, lokalne środowisko, interakcje i reakcje sieci troficznej, a w szczególności antropogeniczne zmiany klimatu i inne przejawy działalności człowieka (Ilustracje 1-3).
Działania człowieka, które bezpośrednio uderzają w mikroorganizmy obejmują emisję gazów cieplarnianych (zwłaszcza CO2, CH4 i N2O), zanieczyszczenie (zwłaszcza eutrofizację), rolnictwo (szczególnie wykorzystywanie gruntów) i wzrost liczebności populacji, który wpływa na zasadzie dodatniego sprzężenia zwrotnego na zmiany klimatu, zanieczyszczenie, praktykę rolniczą oraz rozprzestrzenianie się chorób. Działania człowieka, które zmieniają wskaźniki absorpcji węgla w stosunku do uwalniania wywołują sprzężenie zwrotne dodatnie i zwiększają tempo zmian klimatu. W przeciwieństwie do tego drobnoustroje także zapewniają ważne możliwości rozwiązania problemów powodowanych przez człowieka poprzez lepsze wyniki w zakresie rolnictwa, produkcję biopaliw oraz powstrzymywanie zanieczyszczenia.
Poruszanie konkretnych kwestii związanych z drobnoustrojami wymagać będzie ukierunkowanych badań laboratoryjnych mikrobów modelowych (Ramka 2). Przeprowadzane w laboratorium sondowanie reakcji mikrobiologicznych powinno umożliwić ocenę warunków istotnych z punktu widzenia środowiska i przyjęcie „mikrobocentrycznego” spojrzenia na stresory środowiskowe, a jego kontynuacją powinny być badania w terenie. Eksperymenty dotyczące mezokosmosu oraz badania in situ w terenie są szczególnie ważne dla zyskania wglądu w reakcje na poziomie społeczności na rzeczywiste warunki panujące w środowisku. Skutecznie zaprojektowane badanie wymaga przemyślanego podejmowania decyzji obejmujących wiedzę z licznych dziedzin specyficznych dla biomów morskich (na przykład oceanografia fizyczna) i lądowych (na przykład geochemia).
Aby zrozumieć w jaki sposób różnorodność mikrobiologiczna i funkcjonowanie mikroorganizmów, które kierują wzajemnym oddziaływaniem na niewielką skalę przekładają się na przepływ dużego układu, istotne jest skalowanie wyników od poszczególnych osobników do społeczności i do całych ekosystemów. Zwolennicy modelu systemu ziemskiego muszą wziąć pod uwagę wkład mikrobiologiczny, który wyjaśnia fizjologiczne i adaptacyjne (ewolucyjne) reakcje na przymus biotyczny (uwzględniający inne drobnoustroje, rośliny i służącą jako substrat materię organiczną) i abiotyczny (uwzględniający powierzchnię minerałów, fizykę oceanów oraz chemię).
Musimy poprawić nasze rozumienie ilościowe światowego mikrobiomu morskiego i gleby. Aby zrozumieć obieg biogeochemiczny i sprzężenie zwrotne związane ze zmianami klimatycznymi w dowolnym miejscu na świecie potrzebne nam są informacje ilościowe dotyczące organizmów, które powodują obieg pierwiastków w przyrodzie (w tym ludzi, roślin i mikroorganizmów) oraz warunków środowiskowych (w tym klimatu, właściwości fizjochemicznych gleb, topografii, temperatury oceanów, naświetlenia i mieszania) regulujących aktywność tych organizmów. Struktura modeli ilościowych istnieje, ale modelom tym w dużej mierze brakuje szczegółów mechanistycznych dotyczących drobnoustrojów morskich i lądowych. Przyczyna tej luki ma mniej wspólnego ze sposobem, w jaki konstruuje się taki model matematycznie, ale wywodzi się z braku danych fizjologicznych i ewolucyjnych umożliwiających zdecydowane prognozy reakcji mikrobiologicznych na zmiany klimatu.
Inwestycja ukierunkowana na rozwijanie tej mechanistycznej wiedzy ilustruje ścieżkę krytyczną w stronę generowania globalnych modeli niezbędnych do przeprowadzania testów porównawczych, skalowania i parametryzowania prognoz dotyczących klimatu obecnie i w przyszłości na z wykorzystaniem modelu systemu ziemskiego.
Istniejące obecnie życie ewaluowało przez miliardy lat, czego skutkiem jest olbrzymia bioróżnorodność, zaś różnorodność mikrobiologiczna jest właściwie nieograniczona w porównaniu z życiem makroskopowym. Bioróżnorodność organizmów makroskopowych gwałtownie się zmniejsza na skutek działań człowieka, co wskazuje, że bioróżnorodność mikroorganizmów specyficznych dla żywiciela wśród gatunków zwierzęcych i roślinnych również będzie maleć. Jednakże w porównaniu z organizmami makroskopowymi wiemy znacznie mniej o powiązaniach między drobnoustrojami a antropogenicznymi zmianami klimatu.
Potrafimy rozpoznać wpływ mikrobów na zmiany klimatyczne i zmian klimatycznych na mikroby, ale to, czego się dowiedzieliśmy jest niekompletne, złożone i trudne do zinterpretowania. A zatem nie jest zaskoczeniem fakt, że zdefiniowanie przyczyn i skutków antropogenicznych zmian klimatu na systemy biologiczne stanowi wyzwanie. Pomimo to nie ulega wątpliwości, że działania człowieka powodują zmiany klimatyczne, które zakłócają prawidłowe funkcjonowanie ekosystemu na całym świecie (Ramka 1). W całych biomach morskich i lądowych emisje gazów cieplarnianych wywołane przez mikroby występują coraz częściej i wpływają na zasadzie dodatniego sprzężenia zwrotnego na zmiany klimatu. Bez względu na precyzyjne szczegóły, kompas mikrobiologiczny wskazuje na konieczność podjęcia działań (Ramka 2). Nieznajomość roli, wpływu na zmiany klimatyczne i mechanizmów sprzężenia zwrotnego społeczności mikrobiologicznych w reakcji na te zmiany może sprowadzić niebezpieczeństwo na nas samych. Potrzebne są natychmiastowe, długotrwałe i zdecydowane starania, aby stanowczo włączyć mikroorganizmy do badań, rozwoju technologii oraz decyzji dotyczących polityki i zarządzania. Drobnoustroje nie tylko przyczyniają się do tempa zmian klimatu, ale mogą także wnieść olbrzymi wkład w ich skuteczne łagodzenie i nasze narzędzia adaptacyjne.
Ramka 2 Wezwanie do działania
W swoim ostrzeżeniu mikrobiolodzy apelują o:
-
Większą świadomość, że zdrowie i funkcjonowanie wszystkich organizmów wielokomórkowych, w tym ludzi, zależy od mikroorganizmów; życie mikrobiologiczne stanowi układ wsparcia dla biosfery.
-
Włączenie drobnoustrojów do badań zmian klimatu głównego nurtu, w szczególności do analiz dotyczących przepływu węgla i azotu.
-
Eksperymentalny model, który wyjaśnia zmienne i presje środowiskowe (biotyczne i abiotyczne) istotne dla ekosystemu mikrobiologicznego i reakcji na zmiany klimatyczne.
-
Badanie fizjologicznych, społecznościowych i ewolucyjnych reakcji mikrobiologicznych oraz sprzężenia zwrotnego w odpowiedzi na zmiany klimatu.
-
Skupienie się na mechanizmach mikrobiologicznego sprzężenia zwrotnego podczas monitorowania przepływu gazów cieplarnianych z biomów morskich i lądowych oraz sektora rolniczego, przemysłowego, sektora odpadów i sektora zdrowia, a także o inwestycję w długofalowe obserwacje.
-
Włączenie procesów mikrobiologicznych do modeli ekosystemu i systemu ziemskiego w celu ulepszenia prognoz w obliczu scenariuszy uwzględniających zmiany klimatu.
-
Rozwój innowacyjnych technologii dotyczących mikroorganizmów, aby zminimalizować i złagodzić skutki zmian klimatycznych, zmniejszyć zanieczyszczenie i wyeliminować zależność od paliw kopalnych.
-
Wdrożenie do programów szkolnych nauczania istotnych z punktu widzenia osobistego, społecznego, środowiskowego i pod względem zrównoważonego rozwoju aspektów mikrobiologii z późniejszym rozszerzeniem edukacji w dziedzinie mikrobiologii na poziomie szkolnictwa wyższego, aby uzyskać lepiej wykształcone społeczeństwo oraz właściwie wyszkolonych naukowców i potencjał ludzki.
-
Wyraźne uwzględnienie drobnoustrojów przy podejmowaniu decyzji dotyczących polityki i zarządzania.
-
Uznanie, że wszystkie kluczowe procesy związane z biosferą zależą od mikroorganizmów i znaczny wpływ na nie wywierają działania człowieka, co sprawia, że konieczne staje się włączenie mikrobiologii do zarządzania Celami Zrównoważonego Rozwoju Organizacji Narodów Zjednoczonych i wspierania ich.
Przypisy: